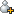圖1 細(xì)菌細(xì)胞從外部環(huán)境中去除Cr(Ⅵ)的可能路徑
Srinath等發(fā)現(xiàn)芽孢桿菌B. circulans,B. megaterium和B.coagulans具有很強(qiáng)的鉻絡(luò)合能力,同時,死細(xì)胞在金屬去除的過程中不受毒性限制。因此與活細(xì)胞相比,死細(xì)胞對鉻的吸附效果更好,并且死細(xì)胞作為生物吸附劑,不需要營養(yǎng)素,吸附的金屬離子易于解吸,生物質(zhì)還可重復(fù)利用。實驗過程中,通常使用熱滅活的細(xì)胞進(jìn)行生物吸附實驗,熱處理導(dǎo)致細(xì)胞壁破裂,釋放更多的官能團(tuán),從而實現(xiàn)生物質(zhì)的最大結(jié)合能力。生物吸附劑對Cr(Ⅵ)的吸附機(jī)理包括在細(xì)菌細(xì)胞壁中金屬與酸性位點(diǎn)之間的離子相互作用和復(fù)合物的形成;胞外多糖以表面黏液層的形式結(jié)合和積累金屬離子,在提高細(xì)胞對重金屬的吸附能力中發(fā)揮了作用。藍(lán)細(xì)菌由于比表面積大,黏液體積大,親和力強(qiáng)和營養(yǎng)要求簡單,因此比其他微生物具有更多的吸附優(yōu)勢。同時發(fā)現(xiàn),隨著水溶液中Cr(Ⅵ)濃度的增加(測試濃度達(dá)20 mg/L),藍(lán)細(xì)菌的金屬去除率增加。據(jù)此推測,由于長期暴露在受重金屬污染的地點(diǎn),這些菌株可能對金屬離子產(chǎn)生了耐受性,顯示了其生物修復(fù)的優(yōu)勢。
1.2細(xì)菌類吸附劑對Cr(Ⅲ)的吸附
多項研究表明:使用細(xì)菌來源生物吸附劑吸附Cr(Ⅲ)時,細(xì)菌合成的胞外多糖不僅為細(xì)菌在極端環(huán)境環(huán)境中提供了保護(hù),而且還提高了其對鉻離子的吸附能力。
發(fā)現(xiàn)Cr(Ⅲ)的吸附與羰基官能團(tuán)密切相關(guān)。Cr(Ⅲ)濃度的增加會促進(jìn)該細(xì)菌產(chǎn)生胞外多糖,提高對Cr(Ⅲ)的生物去除能力。Cr(Ⅲ)的金屬吸附量隨著溶液pH、溫度和初始金屬離子濃度的增加而增加。
2.真 菌
真菌作為生物吸附劑的特征是產(chǎn)生的殘留量小、去除速率快,并且能使工業(yè)發(fā)酵中的真菌廢棄生物質(zhì)增殖。真菌細(xì)胞壁成分在生物吸附中起著重要作用,主要包括纖維素、殼聚糖、甲殼素、糖原、葡聚糖和甘露聚糖等。因此,即使沒有生理活性,真菌生物質(zhì)也可以從水溶液中吸附大量重金屬。
2.1真菌類吸附劑對Cr(Ⅵ)的吸附
在使用毛霉(Mucor hiemalis)、少根根霉(Rhizopus arrhizus)、黑曲霉(Aspergillus niger)、灰黃青霉(Penicillium griseofulvumMSR1)作為生物吸附劑吸附Cr(Ⅵ)時,均發(fā)現(xiàn)Cr(Ⅵ)的去除率隨著pH值的降低而增加,隨著初始Cr(Ⅵ)濃度上升而增加。動力學(xué)研究表明,擬二級模型與實驗數(shù)據(jù)最符合,平衡吸附研究證實Langmuir等溫模型對實驗數(shù)據(jù)的擬合效果最好。此外,從ΔH的值可知,生物吸附過程是吸熱的。
發(fā)現(xiàn)真菌細(xì)胞壁中的氨基、羧酸和巰基參與了鉻的結(jié)合和還原,提出從外部環(huán)境中去除Cr(Ⅵ)的可能路徑:
1)酸性條件下帶正電荷的氨基對帶負(fù)電的重鉻酸根離子的靜電吸附;
2)通過形成瞬態(tài)的Cr(Ⅵ)硫代酯,硫醇將重鉻酸鹽還原為Cr(Ⅲ);
3)硫醇氧化為二硫鍵;
4)Cr(Ⅲ)與氨基配體的螯合作用(圖2)。

圖2 真菌細(xì)胞從外部環(huán)境中去除Cr(Ⅵ)的可能路徑發(fā)現(xiàn)固定化真菌菌絲體比游離真菌菌絲體表現(xiàn)出更高的生物吸附能力和強(qiáng)度。
2.2真菌類吸附劑對Cr(Ⅲ)的吸附
Hanif等利用3種白腐菌(Pleutrotus sajor-cajor、Ganoderma lucidum、Agaricus bitorquis)吸附Cr(Ⅲ),分別進(jìn)行了分批實驗和色譜柱研究,發(fā)現(xiàn)生物吸附量隨著金屬離子濃度(25~200 mg/L)的增加而增加。Langmuir吸附等溫線模型和偽二級動力學(xué)模型能較好地擬合金屬生物吸附數(shù)據(jù),硫酸是從死菌生物質(zhì)中回收金屬的最佳洗脫液。Shoaib等研究少根根霉(Rhizopus arrhizus Fisher)對Cr(Ⅲ)的吸附性能時發(fā)現(xiàn),胺基(—NH2)和羥基(—OH)參與了Cr(Ⅲ)與真菌生物質(zhì)的結(jié)合。
3.藻 類
藻類的生物吸附主要?dú)w因于金屬離子與細(xì)胞壁的結(jié)合,其細(xì)胞壁的主要成分有纖維素、木聚糖、甘露聚糖和海藻酸等,與吸附相關(guān)的基團(tuán)包括羧基、磺酸鹽、羥基和氨基等,它們的相對重要性取決于位點(diǎn)的數(shù)量以及位點(diǎn)與金屬之間的親和力等因素,主要的金屬結(jié)合機(jī)理包括靜電吸引、離子交換和絡(luò)合作用。
3.1藻類吸附劑對Cr(Ⅵ)的吸附
以藻類作為生物吸附劑吸附Cr(Ⅵ)的機(jī)理因使用的藻類不同而存在差異,主要包括靜電吸引和吸附—還原2類鉻吸附機(jī)制。
熱力學(xué)參數(shù)表明,藻類生物對Cr(Ⅵ)的吸附是自發(fā)的、吸熱的。擬一階動力學(xué)模型能很好地描述動力學(xué)數(shù)據(jù),速度控制步驟的過程涉及邊界層以及粒子內(nèi)擴(kuò)散。紅外分析表明—COOH、—OH和—NH2基團(tuán)參與了吸附過程。
Deng等研究剛毛藻(Cladophora albida)對Cr(Ⅵ)的吸附性能時提出其吸附機(jī)制符合吸附-還原機(jī)制,即:
1)Cr(Ⅵ)陰離子與生物質(zhì)表面正電荷基團(tuán)的結(jié)合;
2)相鄰給電子基團(tuán)將Cr(Ⅵ)還原為Cr(Ⅲ);
3)正電荷基團(tuán)與Cr(Ⅲ)離子之間的電子排斥作用使Cr(Ⅲ)離子重新進(jìn)入水相,或Cr(Ⅲ)與相鄰基團(tuán)形成絡(luò)合物。
3.2藻類吸附劑對Cr(Ⅲ)的吸附
水中的Cr(Ⅲ)以陽離子形式存在,這些陽離子與藻類中存在的堿金屬/堿土金屬離子和質(zhì)子發(fā)生離子交換。
Aravindhan等在研究Turbinaria ornate對Cr(Ⅲ)的吸附時發(fā)現(xiàn),其中的羧基與Cr(Ⅲ)絡(luò)合,提出吸收Cr(Ⅲ)的主要官能團(tuán)來自多糖。Li等在研究螺旋藻(Spirulina platensis)對Cr(Ⅲ)的吸附特性時提出:在吸附初期,Cr(Ⅲ)被靜電吸引到藻細(xì)胞表面;然后,通過離子交換與K+、Mn2+、Ca2+、Mg2+、Na+、Fe3+、Zn2+、H+等離子進(jìn)行可逆或不可逆的化學(xué)絡(luò)合作用,吸附的Cr(Ⅲ)大部分與蛋白質(zhì)、多糖、脂類等生物配體穩(wěn)定結(jié)合,而只有很小一部分吸附在藻類表面。Onyancha等也認(rèn)為藻類對Cr(Ⅲ)的吸附是通過離子交換機(jī)制進(jìn)行的,而羧基是主要的結(jié)合位點(diǎn),鉻的配位是通過羰基的氧進(jìn)行的。
4.植 物
植物來源生物吸附劑的優(yōu)點(diǎn)是成本低、來源多種、原料方便、效率相對較高、對環(huán)境的污染很小或幾乎沒有。其中,與吸附相關(guān)的化學(xué)成分包括單寧、木質(zhì)纖維素、多酚和木聚糖等。
4.1植物類吸附劑對Cr(Ⅵ)的吸附
多項研究表明,使用植物來源生物吸附劑吸附Cr(Ⅵ)時,與這些吸附材料結(jié)合的鉻大部分或全部為Cr(Ⅲ)形式。Dupont等研究了從麩皮中提取的木質(zhì)纖維素對Cr(Ⅵ)的去除機(jī)理,發(fā)現(xiàn)吸附反應(yīng)消耗了大量的質(zhì)子,使Cr(Ⅵ)還原為Cr(Ⅲ)。木質(zhì)纖維素部分的氧化與Cr(Ⅵ)還原同時發(fā)生,并導(dǎo)致羥基和羧基的形成,并且羧基的形成有助于增加還原鉻離子交換位點(diǎn)的數(shù)量。而Park等通過XPS光譜發(fā)現(xiàn),含鉻生物材料的光譜與Cr(Ⅲ)的光譜完全匹配,由此推測Cr(Ⅵ)還原為Cr(Ⅲ)的反應(yīng)發(fā)生在材料表面。Suksabye等也發(fā)現(xiàn)大部分結(jié)合在椰殼纖維上的鉻都是Cr(Ⅲ)形式,提出椰殼纖維中木質(zhì)素結(jié)構(gòu)的羰基(—CO)和甲氧基(—O—CH3)基團(tuán)可能與鉻的吸附機(jī)理有關(guān)。椰殼纖維表面上還原的Cr(Ⅲ)可以通過配位共價鍵與—CO基團(tuán)和—O—CH3基團(tuán)鍵合,其中甲氧基和羰基氧原子中的1對孤電子可以與Cr(Ⅲ)形成共價鍵。Vaghetti等在利用巴西松果皮作為生物吸附劑從水溶液中去除Cr(Ⅵ)時提出了多酚對Cr(Ⅵ)的吸附機(jī)制:在酸性條件下重鉻酸鹽將多酚氧化成聚醌,形成的聚醌與重鉻酸鹽還原生成的Cr(Ⅲ)形成絡(luò)合物。
4.2植物類吸附劑對Cr(Ⅲ)的吸附
使用植物來源生物吸附劑吸附Cr(Ⅲ)是一個快速的過程,其機(jī)理包括靜電吸引和絡(luò)合作用等。
Chojnacka用麥秸稈作為生物吸附劑吸附Cr(Ⅲ),發(fā)現(xiàn)生物吸附在10~20 min內(nèi)達(dá)到平衡。并且由于麥秸稈通常含量豐富且成本低,因此可以通過增加吸附劑的濃度來提高廢水處理效率。吸附過程的動力學(xué)可以用擬二級方程描述,生物吸附平衡可以用Freundlich方程描述,其生物吸附機(jī)理為物理吸附。Flogeac等進(jìn)一步研究了從麥秸中提取的木質(zhì)纖維素與Cr(Ⅲ)的結(jié)合能力,EPR(電子順磁共振波譜)和XAS(X射線吸收光譜)的微觀研究表明,Cr(Ⅲ)在木質(zhì)纖維素上的吸附是通過形成內(nèi)表面復(fù)合物來實現(xiàn)的。EXAFS(X射線吸收精細(xì)結(jié)構(gòu))研究得出了金屬周圍精確的局部結(jié)構(gòu)——八面體構(gòu)型。XANES(X射線吸收近邊緣結(jié)構(gòu))光譜表明,Cr(Ⅲ)與木質(zhì)纖維素的羧基結(jié)合。
5.其 他
除了以上提到的細(xì)菌、真菌、藻類和植物來源的天然生物吸附劑以外,為了提高材料對鉻離子的吸附性能,大量經(jīng)天然生物吸附材料改性制成的吸附劑亦被應(yīng)用。Bingol等先后研究了陽離子表面活性劑修飾的酵母、地衣從水溶液中去除鉻酸根陰離子的能力,發(fā)現(xiàn)用十六烷基三甲基溴化銨(CTAB)進(jìn)行改性,可大大改善生物吸附效率。
而植物來源的生物吸附劑中,大多含有木質(zhì)素,其結(jié)構(gòu)中富含醇羥基和酚羥基,在吸附過程中起著重要作用。也因此,木質(zhì)素的改性集中在酯化和醚化反應(yīng)上。據(jù)此,Wang等提出通過以下3種方法對木質(zhì)素進(jìn)行改性,以提高對鉻的吸附效率:
1)將木質(zhì)素部分解聚或斷裂成小片段或芳香大分子單體;
2)通過其他溶劑產(chǎn)生新的化學(xué)活性位;
3)酚和醇羥基的化學(xué)修飾,以獲得木質(zhì)素基吸附材料的大表面積,強(qiáng)結(jié)合位點(diǎn)以及較好的吸附性能。
Yao等發(fā)現(xiàn),枯草芽孢桿菌NX-2衍生的聚谷氨酸(γ-PGA)具有配體功能,通過游離α-羧基與金屬離子配位,達(dá)到去除Cr(Ⅲ)的效果。除此之外,經(jīng)過HAc溶解、NaOH沉淀等步驟改性而成的殼聚糖微珠、用殼聚糖與Ustilago(真菌孢子)制成的復(fù)合微膠囊、以及利用簡單化學(xué)處理改性的稻草作為生物吸附劑吸附Cr(Ⅲ),均被證實有良好效果。
03展 望
利用生物吸附法修復(fù)鉻污染場地一直以來都是研究者不斷探索的領(lǐng)域,已有大量文獻(xiàn)報道了使用細(xì)菌、真菌、藻類和植物來源的生物吸附劑從土壤、水體中去除鉻的方法,許多生物質(zhì)材料在實驗室條件下表現(xiàn)出良好的吸附性能,且其低成本、不會造成二次污染以及操作簡單的特點(diǎn),使其在眾多重金屬修復(fù)方法中具有一定優(yōu)勢。但同時生物吸附修復(fù)技術(shù)仍然存在一些缺點(diǎn),比如活生物質(zhì)對鉻的耐受能力有限,以其作為生物吸附劑時,僅適用于一些輕度或中度的鉻污染場地。除此之外,生物吸附的過程還容易受到外部環(huán)境的干擾。
因此,為了充分利用生物吸附劑修復(fù)鉻污染場地,首先需要闡明微生物代謝的理論機(jī)制,以便篩選更多的耐鉻微生物菌株,并分析微生物菌株在重金屬修復(fù)中的安全性和可控性。同時,鉻污染生物吸附修復(fù)的實驗室研究表明,吸附過程受到溫度、pH、其他金屬離子等各種因素的影響,而在實際場地應(yīng)用中,吸附過程所面臨的環(huán)境更為復(fù)雜易變。因此需要對污染場地進(jìn)行長期調(diào)查。其次,就目前的研究來說,所涉及的吸附過程和機(jī)理仍存在一些不確定性,鉻修復(fù)過程的機(jī)理闡釋仍需大量努力和實踐。最后,大量文獻(xiàn)報道證實改性天然生物質(zhì)材料是提高吸附性能的有效手段。材料改性通過引入更多的功能性活性基團(tuán),且通過增加表面積、化學(xué)穩(wěn)定性和機(jī)械強(qiáng)度,來提升吸附劑的吸附能力。同時,改性材料更易于實現(xiàn)循環(huán)利用,提升吸附材料的經(jīng)濟(jì)性并降低對環(huán)境的影響。